The prokaryotes, which include Bacteria and Archaea, are mostly single-celled organisms that, by definition, lack membrane-bound nuclei and other organelles. A bacterial chromosome is a closed circle that, unlike eukaryotic chromosomes, is not organized around histone proteins. The central region of the cell in which prokaryotic DNA resides is called the nucleoid region. In addition, prokaryotes often have abundant plasmids, which are shorter, circular DNA molecules that may only contain one or a few genes. Plasmids can be transferred independently of the bacterial chromosome during cell division and often carry traits such as those involved with antibiotic resistance.
Transcription in prokaryotes (and in eukaryotes) requires the DNA double helix to partially unwind in the region of mRNA synthesis. The region of unwinding is called a transcription bubble. Transcription always proceeds from the same DNA strand for each gene, which is called the template strand. The mRNA product is complementary to the template strand and is almost identical to the other DNA strand, called the nontemplate strand, or the coding strand. The only nucleotide difference is that in mRNA, all of the T nucleotides are replaced with U nucleotides (Figure). In an RNA double helix, A can bind U via two hydrogen bonds, just as in A–T pairing in a DNA double helix.
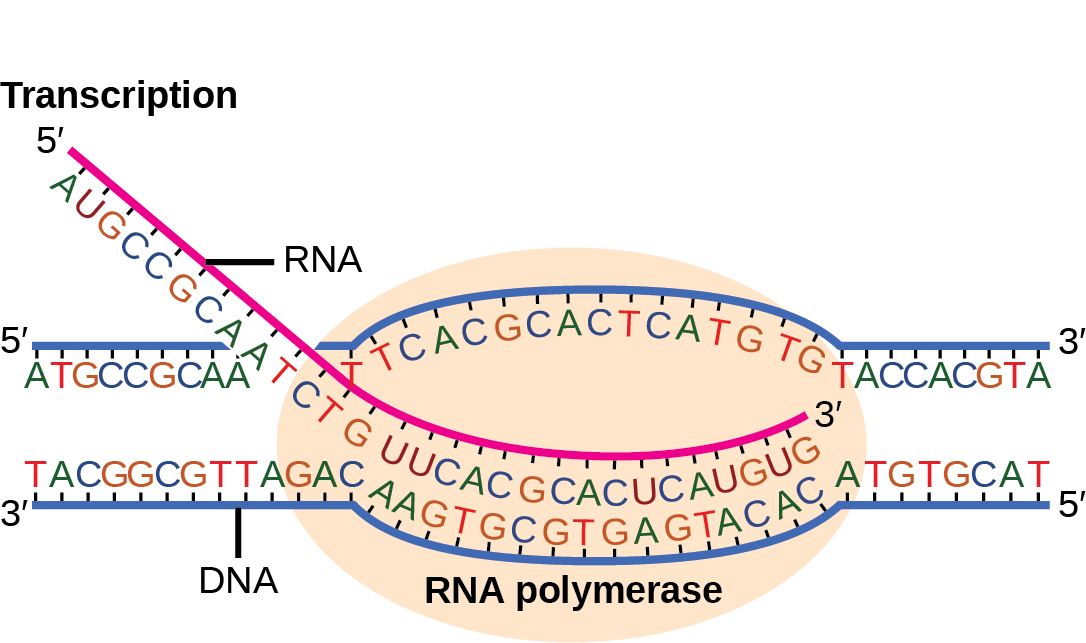
The nucleotide pair in the DNA double helix that corresponds to the site from which the first 5' mRNA nucleotide is transcribed is called the +1 site, or the initiation site. Nucleotides preceding the initiation site are denoted with a “-” and are designated upstream nucleotides. Conversely, nucleotides following the initiation site are denoted with “+” numbering and are called downstream nucleotides.